Vadim Sharov
Vadim Sharov

Великий шведский биолог Карл Линней разделил весь природный мир на три царства: минералы, растений и животных; отношения между отдельными видами он не учитывал. Столетие спустя Дарвин опубликовал «Происхождение видов», и картина начала усложняться. Наконец, в 1969 г. в журналеScience вышла эколога Роберта Уиттекера, и его концепция царств живой природы на полвека стала канонической: простейшие прокариоты, затем одноклеточные эукариоты, а надо всем этим царства растений, грибов и животных. Но теперь генетические исследования поставили ее под сомнение.
От Линнея до Уиттекера
Как известно, основу современной систематики живых существ заложил великий шведский биолог Карл Линней. В своем эпохальном труде «Система Природы» (Systema Naturae, 1735) он разделил весь природный мир на три царства: минералы, растений и животных. Отношения между отдельными видами - то, как они происходили друг от друга, конкурировали, появлялись и исчезали - при этом не брались в расчет: Линней хотел всего лишь создать упорядоченную классификацию «божьих творений». До выхода «Происхождения видов» Дарвина оставалось еще 124 года.
После торжества дарвиновской теории и по мере того, как накапливались все новые знания о живых существах, картина начала усложнятся. Стало ясно, что грибы стоят особняком от животных и растений. Кроме того, были изучены одноклеточные животные - как имеющие в своих клетках ядро (эукариоты), так и лишенные его (прокариоты). Всем им тоже нужно было найти места на эволюционном древе.
В результате, в 1969 г. эколог Роберт Уиттекер (Robert Whittaker), обобщив и несколько упростив собранные своими предшественниками данные, предложил в своей статье в журналеScience новую концепцию царств живой природы (камни биологов, естественно, уже не интересовали):

Как мы видим, на схеме Уиттекера основание «ствола» эволюционного древа представляют собой простейшие прокариоты (Monera ), верхушку - царство одноклеточных эукариот (Protista ), а венчают всю конструкцию три мощных ветви - царства растений (Plantae ), грибов (Fungi ) и животных (Animalia ). Именно эта схема стала канонической на многие десятилетия и вошла в учебники, по которым учились мы с вами.
Экология и генетика
Схема Уиттекера не случайно снискала такое широкое признание - она проста, удобна и, как тогда казалось, хорошо согласуется с фундаментальными принципами существования живой материи.
Например, с точки зрения экологии (мы не зря упомянули, что Уиттекер был экологом) все живые существа по типу питания делятся на три типа. Растения-продуценты занимаются тем, что продуцируют, то есть создают биомассу с помощью фотосинтеза, вдыхая на солнечном свету углекислый газ и выделяя кислород. Животные-консументы не создают, а только потребляют эту созданную растениями биомассу. Причем собственно траву едят только консументы 1-го порядка (травоядные), консументы 2-го порядка (хищники) едят консументов 1-го порядка, и так далее, выше по пищевой цепочке. Наконец, грибы - редуценты, они разлагают биомассу на более простые соединения, чтобы снова включить ее в круговорот. Простое эволюционное древо как раз и отражало историческое разделение живых существ в соответствии с этими тремя стратегиями.
Первые данные генетических исследований тоже, казалось бы, подтверждали правильность схемы Уиттекера. Сравнение вариаций одного гена, кодирующего одну из частей рибосомной РНК у разных живых существ давало схожую картину с тремя мощными ветвями - растениями, животными и грибами - наверху и одноклеточными внизу.
Но что-то пошло не так…
Однако генетический анализ показал другое. Оказалось, что некоторые митохондриальные гены присутствуют в клетках лямблий и трихомонад. Посмотрев внимательнее в электронные микроскопы, исследователи заметили там и рудименты самих митохондрий - крошечные редуцированные пузырьки, которые невозможно заметить, если не знаешь, что искать.
Одним словом, выяснилось, что старое эволюционное древо «по Уиттекеру» ложно трактует родственные отношения между живыми существами, которые на проверку оказались гораздо более сложными. Чтобы отразить современные уточненные представления об этом, биологи недавно «нарисовали» новое, более сложное эволюционное древо, в котором вместо царств главную роль играют супергруппы, а животные (включая людей) оказываются ближе к одноклеточным хоанофлагеллятам, чем к другим многоклеточным организмам. Об этой новой схеме мы расскажем во второй части статьи.
Vadim Sharov
Новое уточненное эволюционное древо животного царства, основанное на анализе рекордного числа генов и типов животных, позволило разрешить многие спорные вопросы эволюции и систематики. Подтвердилась теория, согласно которой разделение на первичноротых и вторичноротых произошло еще до того, как у животных сформировался цело м (вторичная полость тела). Первичноротые подразделяются на две четкие эволюционные линии: Lophotrochozoa (плоские и кольчатые черви, моллюски, брахиоподы, немертины) и Ecdysozoa (круглые и головохоботные черви, членистоногие, онихофоры, тихоходки).
Вплоть до последней четверти XX века биологи реконструировали эволюционную историю животных преимущественно на основе данных сравнительной анатомии, эмбриологии и палеонтологии. Затем к этому списку добавились молекулярные данные, самыми важными из которых являются нуклеотидные последовательности ДНК. Эволюционные реконструкции («деревья»), основанные на молекулярных данных, не всегда совпадали со старыми «классическими» деревьями. Это приводило к бурным спорам среди зоологов.
Поначалу многие придерживались мнения, что старые проверенные методы надежнее новомодных молекулярных. Но постепенно чаша весов склонилась в другую сторону, и сегодня большинство специалистов считает, что молекулярные данные в принципе позволяют гораздо более точно реконструировать пути эволюции животных, чем морфологические и эмбриологические признаки. В России, правда, до сих пор многие не согласны с этим, но на Западе таких «ретроградов», не доверяющих молекулярным реконструкциям, осталось очень мало.
Молекулярные «признаки» (последовательности нуклеотидов) имеют два важных преимущества по сравнению с морфологическими. Во-первых, их просто гораздо больше. Фактически каждый нуклеотид в хромосоме можно рассматривать как отдельный признак - и таким образом получать деревья, основанные на многих сотнях и тысячах признаков, тогда как число морфологических признаков, пригодных для филогенетического (эволюционного) анализа, обычно ограничивается несколькими десятками. Во-вторых, большинство морфологических признаков непосредственно влияют на жизнеспособность организма, тогда как замены многих нуклеотидов являются нейтральными (безразличными). Морфологическое сходство не обязательно свидетельствует о родстве - оно может развиться и у неродственных организмов под воздействием естественного отбора в сходных условиях обитания (это явление называют конвергенцией). Конвергентное возникновение сходных нуклеотидных последовательностей гораздо менее вероятно.
Проблема, однако, состоит в том, что достоверность любых эволюционных реконструкций, в том числе молекулярных, очень сильно зависит от объема и полноты исходных данных.
Главным критерием достоверности молекулярных деревьев считается их устойчивость или повторяемость. Существует несколько разных алгоритмов построения дерева на основе одного и того же массива исходных данных (например, нуклеотидных последовательностей какого-нибудь гена у нескольких разных организмов). Если применение разных алгоритмов дает одинаковый результат, это свидетельствует о его надежности. Разработаны также специальные процедуры для оценки достоверности «узлов» (точек ветвления) получающихся деревьев (см.: bootstrapping).
Первые молекулярные деревья животного царства, основанные на единичных генах и очень небольшом количестве видов, отличались низкой устойчивостью, и потому вызывали мало доверия. Очень скоро стало ясно, что чем больше генов и групп животных вовлечено в анализ, тем устойчивее и надежнее становятся результаты. Ученые, разумеется, стали изо всех сил наращивать объем используемых данных. Постепенно стала вырисовываться картина, довольно сильно отличающаяся от «классической», основанной на морфологии и эмбриологии.
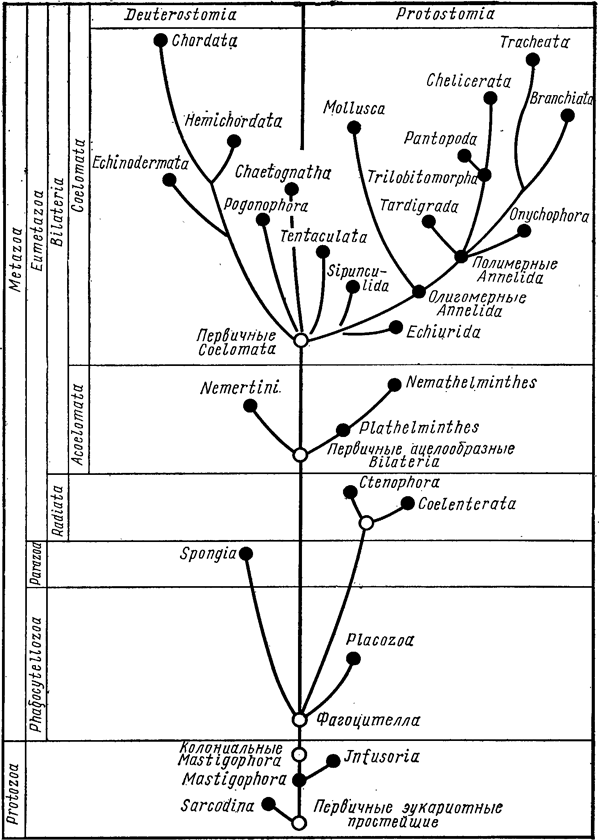
Важнейшее из выявленных отличий касалось родственных отношений между основными типами двусторонне-симметричных животных (билатерий). Согласно классическим представлениям, все билатерии, имеющие целом (вторичную полость тела), происходят от общего предка и противопоставляются «доцеломическим» билатериям, таким как плоские и круглые черви. Целоматы подразделяются на первичноротых (кольчатые черви, моллюски, членистоногие и др.) и вторичноротых (хордовые, полухордовые, иглокожие). Кольчатые черви считались предками членистоногих.
Молекулярные данные, напротив, показывали, что разделение на две линии, соответствующие первично- и вторичноротым, произошло раньше, еще до того, как у билатерий появился цело м. Из этого следовало, что цело м, который специалисты по сравнительной анатомии считали надежнейшим таксономическим признаком (основой для естественной классификации), в действительности развился независимо у первичноротых и вторичноротых. Не имеющие целома круглые черви, согласно молекулярным данным, оказались близкими родственниками членистоногих (их объединили в группу «линяющих» - Ecdysozoa), а плоские черви - родней моллюсков, а также кольчатых червей, родство которых с членистоногими молекулярные данные никак не хотели подтверждать. Плоских червей (не имеющих целома), а также имеющих цело м моллюсков, кольчатых червей и ряд других типов объединили в группу Lophotrochozoa.
Все эти выводы, однако, до самого последнего времени нельзя было признать окончательными. Молекулярные деревья оставались неустойчивыми. Некоторые из них как будто подтверждали «старую» версию эволюции животных, в которой целом появился лишь однажды (эта версия получила название «целоматной теории»). Чтобы разрешить это противоречие, исследователи постепенно наращивали объем и представительность используемых молекулярных данных в надежде, что получаемые деревья в конце концов обретут устойчивость.
Группа ученых из США, Дании, Германии и Великобритании опубликовала в последнем номере журнала Nature новейший вариант молекулярного эволюционного древа животных, основанный на рекордном количестве генов (150) и групп животных. В анализе использовано 77 видов, относящихся к 21 типу животных, причем для 11 из этих типов геномные данные до сих пор отсутствовали. Многие части (узлы) получившегося дерева действительно оказались значительно более устойчивыми, чем в более ранних исследованиях такого рода.

Полученные результаты убедительно свидетельствуют против классической «целоматной теории». Самыми «примитивными» из включенных в анализ групп оказались гребневики. Билатерии сначала подразделяются на линии первично- и вторичноротых, и только потом в каждой из этих линий независимо формируется цело м. Первичноротые подразделяются на Lophotrochozoa и Ecdysozoa. Ближайшими родственниками членистоногих оказались онихофоры и тихоходки (что соответствует классическим представлениям), а также круглые черви (что совершенно не соответствует оным). Ближайшими родственниками кольчатых червей оказались не членистоногие, как считалось ранее, а брахиоподы и немертины.
Многое прояснилось, однако родственные связи некоторых групп так и остались неопределенными (их положение на новом древе оказалось неустойчивым). Эти группы не показаны на приведенном рисунке (за исключением губок). Причины неустойчивости авторы видят в том, что для одних групп они не сумели собрать достаточное количество молекулярных данных (губки, бескишечные турбеллярии, мизостомиды), а другие были представлены недостаточным количеством видов (мшанки, коловратки). Кроме того, авторы не смогли включить в анализ трихоплакса, который, судя по результатам анализа митохондриальной ДНК, является самым примитивным из современных животных.
Эукариотические организмы, специализирующиеся на гетеротрофном питании, дали начало Животным и Грибам.
В протерозойской эре возникают все известные типы Многоклеточных беспозвоночных животных. Существует две основные теории происхождения многоклеточных животных. Согласно теории гастреи (Э. Геккель), исходным способом формирования двуслойного зародыша является инвагинация (впячивание стенки бластулы). Согласно теории фагоцителлы (И. И. Мечников), исходным способом формирования двуслойного зародыша является иммиграция (перемещение отдельных бластомеров в полость бластулы). Возможно, эти две теории взаимно дополняют друг друга.
Кишечнополостные – представители наиболее примитивных (двуслойных) многоклеточных: их тело состоит всего из двух слоев клеток: эктодермы и энтодермы. Уровень дифференцировки тканей очень низкий.
У Низших червей (Плоские и Круглые черви) появляется третий зародышевый листок – мезодерма. Это крупный ароморфоз, благодаря которому появляются дифференцированные ткани и системы органов.
Затем эволюционное древо животных разветвляется на Первичноротых и Вторичноротых. Среди Первичноротых у Кольчатых червей образуется вторичная полость тела (целом). Это крупный ароморфоз, благодаря которому становится возможным разделение тела на отделы.
Кольчатые черви имеют примитивные конечности (параподии) и гомономную (равнозначную) сегментацию тела. Но в начале кембрия появляются Членистоногие, у которых параподии преобразованы в членистые конечности. У Членистоногих появляется гетерономная (неравнозначная) сегментация туловища. У них имеется хитиновый наружный скелет, который способствует появлению дифференцированных пучков мышц. Перечисленные особенности Членистоногих являются ароморфозами.
Наиболее примитивные Членистоногие – Трилобитообразные – господствовали в палеозойских морях. Современные Жабродышащие первичноводные членистоногие представлены Ракообразными. Однако в начале девона (после выхода на сушу растений и формирования наземных экосистем) происходит выход на сушу Паукообразных и Насекомых.
Насекомые наиболее приспособлены к жизни на суше, благодаря появлению крупных ароморфозов:
– Наличие зародышевых оболочек – серозной и амниотической.
– Наличие крыльев.
– Пластичность ротового аппарата.
С появлением Цветковых растений в меловом периоде начинается совместная эволюция Насекомых и Цветковых (коэволюция), и у них формируются совместные адаптации (коадаптации). В кайнозойской эре Насекомые, как и Цветковые растения, находятся в состоянии биологического прогресса.
Среди Вторичноротых животных наивысшего расцвета достигают Хордовые животные, у которых появляется ряд крупных ароморфозов: хорда, нервная трубка, брюшная аорта (а затем – сердце).
От примитивных Хордовых животных в силуре происходят первые Позвоночные (Бесчелюстные). У позвоночных формируется осевой и висцеральный скелет, в частности, мозговая коробка и челюстной отдел черепа, что также является ароморфозом. Низшие Челюстноротые позвоночные представлены разнообразными Рыбами. Современные классы рыб (Хрящевые и Костные) формируются в конце палеозоя – начале мезозоя).
Часть Костных рыб (Мясистолопастные), благодаря двум ароморфозам – легочному дыханию и появлению настоящих конечностей – дала начало первым Четвероногим – Амфибиям (Земноводным). Первые Земноводные вышли на сушу в девонском периоде, но их расцвет приходится на каменноугольный период (многочисленные стегоцефалы). Современные Амфибии появляются в конце юрского периода.
Параллельно среди Четвероногих появляются организмы с зародышевыми оболочками – Амниоты. Наличие зародышевых оболочек – крупный ароморфоз, который впервые появляется у Рептилий. Благодаря зародышевым оболочкам, а также ряду других признаков (ороговевающий эпителий, тазовые почки, появление коры больших полушарий) Рептилии полностью утратили зависимость от воды. Появление первых примитивных рептилий – котилозавров – относится к концу каменноугольного периода. В перми появляются разнообразные группы рептилий: зверозубые, первоящеры и другие. В начале мезозоя формируются ветви черепах, плезиозавров, ихтиозавров. Начинается расцвет рептилий.
От групп, близких к первоящерам, отделяются две ветви эволюционного развития. Одна ветвь в начале мезозоя дала начало многочисленной группе псевдозухий. Псевдозухии дали начало нескольким группам: крокодилы, птерозавры, предки птиц и динозавры, представленные двумя ветвями: ящеротазовые (бронтозавр, диплодок) и птицетазовые (только растительноядные виды – стегозавр, трицератопс). Вторая ветвь в начале мелового периода привела к появлению подкласса чешуйчатых (ящерицы, хамелеоны и змеи).
Однако Рептилии не смогли утратить зависимость от низких температур: теплокровность у них невозможна из-за неполного разделения крови на венозную и артериальную. В конце мезозоя с изменением климата происходит массовое вымирание рептилий.
Лишь у части псевдозухий в юрском периоде появляется полная перегородка между желудочками, редуцируется левая дуга аорты, происходит полное разделение кругов кровообращения, и становится возможной теплокровность. В дальнейшем эти животные приобрели ряд адаптаций к полету и дали начало классу Птицы.
В юрских отложениях мезозойской эры (≈ 150 млн. лет назад) обнаружены отпечатки Первоптиц: археоптерикса и археорниса (три скелета и одно перо). Вероятно, это были древесно-лазающие животные, которые могли планировать, но не были способны к активному полету. Еще раньше (в конце триаса, ≈ 225 млн. лет назад) существовал протоавис (два скелета обнаружены в 1986 году в Техасе). Скелет протоависа существенно отличался от скелета рептилий, большие полушария мозга и мозжечок были увеличены в размерах. В меловом периоде существовали две группы ископаемых птиц: ихтиорнисы и гесперорнисы. Современные группы птиц появляются только в начале кайнозойской эры.
Существенным ароморфозом в эволюции птиц можно считать появление четырехкамерного сердца в сочетании с редукцией левой дуги аорты. Произошло полное разделение артериальной и венозной крови, что сделало возможным дальнейшее развитие головного мозга и резкое повышение уровня обмена веществ. Расцвет Птиц в кайнозойской эре связан с рядом крупных идиоадаптаций (появление перьевого покрова, специализация опорно-двигательного аппарата, развитие нервной системы, забота о потомстве и способность к перелетам), а также с рядом признаков частичной дегенерации (например, утрата зубов).
В начале мезозойской эры появляются первые Млекопитающие, которые возникли благодаря целому ряду ароморфозов: увеличенные полушария переднего мозга с развитой корой, четырехкамерное сердце, редукция правой дуги аорты, преобразование подвеска, квадратной и сочленовой костей в слуховые косточки, появление шерстного покрова, млечных желез, дифференцированных зубов в альвеолах, предротовой полости.
В юрском периоде мезозойской эры Млекопитающие были представлены, как минимум, пятью классами (Многобугорчатые, Трехбугорчатые, Трикодонты, Симметродонты, Пантотерии). Один из этих классов, вероятно, дал начало современным Первозверям, а другой – Сумчатым и Плацентарным. Плацентарные млекопитающие, благодаря появлению плаценты и настоящего живорождения, в кайнозойской эре переходят в состояние биологического прогресса.
Исходным отрядом Плацентарных являются Насекомоядные. От Насекомоядных рано отделились Неполнозубые, Грызуны, Приматы и ныне вымершая группа Креодонтов – примитивных хищников. От Креодонтов отделились две ветви. Одна из этих ветвей дала начало современным Хищным, от которых отделились Ластоногие и Китообразные. Другая ветвь дала начало примитивным копытным (Кондилартрам), а затем Непарнокопытным, Парнокопытным и родственным отрядам.
Окончательная дифференцировка современных групп Млекопитающих завершилась в эпоху великих оледенений – в плейстоцене. На современный видовой состав Млекопитающих значительное влияние оказывает антропогенный фактор. В историческое время были истреблены: тур, стеллерова корова, тарпан и другие виды.
В конце кайнозойской эры у части Приматов возникает особый тип ароморфоза – переразвитие коры больших полушарий головного мозга. В результате возникает совершенно новый вид организмов – Человек разумный.
Царство грибов совмещает черты строения растений и животных, иначе говоря, это самостоятельное царство эукариотов – гетеротрофов.
Лишайники – симбиотические организмы, состоящие из двух компонентов: водоросль и гриб. Лишайники бывают накипные, листоватые и кустистые.
Царство растений включает низшие растения (водоросли) и высшие растения (все остальные группы).
Водоросли произошли от способных к фотосинтезу прокариотов, т.е. синезеленых водорослей (цианей). Одноклеточные эукариотические водоросли дали начало многоклеточным водорослям (бурые, красные, зеленые, золотистые). Многоклеточные водоросли дали начало псилофитам, а они, в свою очередь, мхам. Моховидные – это обособленная и тупиковая ветвь развития растения. От псилофитов произошли плауновидные, хвощевидные и папоротниковидные. От первичных разноспоровых папоротниковых возникли голосеменные. Представители современных голосеменных растений; гинкго, сосна, ель, пихта, лиственница, кедр, кипарис, можжевельник, гнетум, эфедра, саговник). Наиболее современная и многочисленная группа покрытосеменных эволюционировала параллельно голосеменным от общего с ним предка семенного папоротника. Представители покрытосеменных – это двудольные и однодольные.
Эволюционное древо животных
Царство животных подразделяют на два подцарства: одноклеточные и многоклеточные.
Одноклеточные организмы (эукариоты) произошли от гетеротрофных прокариотов. В современной фауне к ним относят корненожек, жгутиковых, споровиков, инфузорий.
Дальнейшее развитие происходит от примитивных турбеллярий с образованием кольчатых червей (малощетинковые, пиявки, многощетинковые). Примитивные многощетинковые черви определяют возникновение четырех ветвей в древе животных.
Первая ветвь – моллюски (брюхоногие, двустворчатые, головоногие).
Вторая ветвь – членистоногие (ракообразные, паукообразные, насекомые).
Третья ветвь – иглокожие (морские звезды, морские ежи и голотурии, или морские огурцы).
Четвертая ветвь – хордовые, которые возникают вначале палеозоя, когда все типы беспозвоночных животных (рассмотренных выше) уже существовали. Произошли хордовые животные от общего с иглокожими вторичноротого двустороннесимметричного свободноплавающего предка.
Тип хордовых объединяет 3 крупные группы животных: подтипы бесчерепных, личиночнохордовых и черепных, или позвоночных. Подтип бесчерепные состоит из одного класса животных – головохордовых, всего их 30 видов, например, ланцетник. Подтип личиночнохордовые (или оболочники) произошли от примитивынх свободноплавающих бесчерепных, которые перешли к сидячему образу жизни. Оболочники все являются морскими организмами, среди наиболее известных – асцидии.
Высшим подтипом хордовых являются позвоночные. Среди позвоночных выделяют круглоротых (бесчелюстных) – это миноги, миксины. От примитивных круглоротых произошли рыбы, которые делятся на хрящевые, костные, кистеперые, двоякодышащие. Кистеперые рыбы дали начало земноводным, или амфибиям. Амфибии включают в себя хвостатых, бесхвостых, безногих. Например, протеи, тритоны, саламандры и сирены; жабы и лягушки; рыбозмеи и червяги. От амфибий произошли рептилии, или пресмыкающие. В современной фауне присутствуют отряды чешуйчатых (змеи, ящерицы, двуходки, хамелеоны), крокодилов, черепах и клювоголовых (гаттерии).
От неспециализированных, лазающих пресмыкающих произошли птицы. Современные птицы включают группы килевых, или летающих; плавающих, или пингвины; бескилевых, или бегающие (страусы, киви, казуары).
Предками млекопитающих являются неспециализированные палеозойские рептилии с чертами строения земноводных, или зверозубые рептилии. Первые млекопитающие дивергировали на две ветви. Первая ветвь – это первозвери (однопроходные), например, ехидна, утконос. Вторая ветвь – это сумчатые (коала, кенгуру, опоссумы), а также плацентарные (землеройки, летучие мыши, грызуны, хищные, ластоногие, парнокопытные, непарнокопытные, слоны, приматы, человек). Линия человека начинает развиваться от предковых форм насекомоядных полуобезьян.
Родословное древо (филема) -способ изображения родственных связей и эволюции организмов в виде дерева. Еще Ч. Дарвин в 1859 г. писал, что «это сравнение очень близко соответствует истине. Зеленые ветви с распускающимися почками представляют существующие виды, а ветви предшествующих лет соответствуют длинному ряду вымерших видов». Сам Дарвин дал в 1859 г. лишь схему возникновения многих видов, родов и семейств от одного общего вида-предка, но уже в 1866 г. Э. Геккель изобразил первое родословное древо всех живых существ (см. рис.). Геккель различал три основных царства живой природы - растений, протистов и животных.
Так представлял немецкий ученый XIX в. Э. Геккель родословную животных, которую он как зоолог разработал наиболее подробно и близко к действительности.
Его древо отражало уровень биологических знаний того времени, он сближал далекие по современным представлениям группы. Сейчас биологи склоняются к мысли, что жизнь вскоре после ее возникновения расщепилась на три ствола, которые называют надцарствами. Два из них известны уже давно - это организмы, не имеющие оформленного ядра (прокариоты), и ядерные организмы (эукариоты). Сравнительно недавно некоторые систематики прокариот стали разделять на два самостоятельных надцарства - настоящих бактерий (эубактерий) и архебактерий. По некоторым чертам строения и обмена веществ архебактерий оказываются близки к эукариотам. По-видимому, архебактерий сохранили больше, чем другие организмы, черты исходного праорганизма.
Эубактерий включают бактерий и группу, называвшуюся ранее синезелеными водорослями (цианобактерии). Построить их родословное древо удалось лишь в последние годы, использовав сравнительные данные по строению их рибосомных РНК.
По-видимому, ветви архебактерий, эубактерий и предков организмов с оформленным ядром - эукариот разошлись от общего ствола жизни практически одновременно.
Дальнейшая история эукариот связана, вероятно, с симбиозом - какие-то аэробные бактерии стали обитать в цитоплазме их клеток. Так могли возникнуть митохондрии. С тех пор жизнь эукариот неразрывно связана с аэробным, кислородным дыханием, лишь немногие многоядерные амебы, обитающие в бескислородных илах, потеряли его уже вторично.
Впрочем, эту теорию симбиозогенеза эукариот разделяют не все.
Второй этап симбиоза: внедрение в эукариотические клетки каких-то синезеленых организмов - предков хлоропластов - привел к возникновению хлорофиллоносных организмов - растений. Сначала это были одноклеточные зеленые водоросли, но из них возникло все разнообразие современной флоры.
Надцарство эукариот теперь обычно разделяют на три ветви - три царства - животных, растений и грибов. Но не все в этой схеме ясно. Загадочные слизевики, например, настолько далеко отстоят от всех трех царств, что, похоже, заслуживают выделения в четвертое. Споры идут и о месте в филеме простейших, одноклеточных эукариот. Ведь одни из них ближе к растениям (эвглена, вольвокс и др.), другие - к животным. Но выделить простейших в самостоятельную ветвь, как это сделал Геккель, вряд ли возможно. Слишком они разнообразны. Современные составители древ колеблются - разделить простейших по трем основным царствам эукариот или же создавать новые царства. Число основных ветвей ядерных организмов тогда возрастет чуть ли не до десятка.
Основные ветви-царства в родословном древе делятся на более мелкие - типы. О числе этих мелких ветвей, порядке их расположения, а также о времени ответвления до сих пор идут ожесточенные споры. Одних животных систематики насчитывают до 33 типов. Не все из них имели одну эволюционную судьбу: в кроне «древа жизни» имеются пышно разветвленные побеги вроде огромных типов членистоногих, моллюсков или хордовых и тоненькие веточки, представленные немногими десятками видов. Но все они в равной степени интересны систематикам-эволюционистам. Ведь родословные древа - наглядное изображение процесса филогенеза.
В настоящее время родословные древа строятся не только на основании данных морфологии, эмбриологии и палеонтологии, как во времена Геккеля и в последующие годы. Для сравнения используют данные молекулярной биологии о последовательности аминокислот в белках и нуклеотидов в РНК и ДНК. Для сравнения внутри относительно небольших и не очень древних групп, таких, как позвоночные, используют быстро меняющиеся в эволюции белки, например гемоглобин. Для анализа же событий, происшедших миллиарды лет назад, используют такие мало меняющиеся (консервативные) молекулы, как рибосомные РНК.
